脳が対側支配をする進化的な利点は何か
- 正中を横切る神経線維
- 神経線維の交叉に関連した疾患
- Ramón y Cajalの仮説
- ねじれ仮説 (twist hypothesis)
- 回避行動仮説 (avoidance behavior hypothesis)
- 保護機構仮説 (protective mechanism hypothesis)
- まとめ
- 参考文献
神経解剖を学ぶと、次のような疑問は自然と生じると思います。それは「なぜ脳では対側支配(contralateral innervation)があるのか」というものです。ここでの対側とは、身体の正中線に対し反対側のことを指します。例としては左運動野が右側の筋群を、右運動野 が 左側の筋群 を制御するといった対側制御(contralateral control)が挙げられます。何ともややこしい配線ですが、なぜ同側支配ではダメだったのでしょうか?対側支配をする利点はあるのでしょうか?
かなり基本的な内容に対する疑問ではありますが、この問題はRamón y Cajalの1898年の論文に端を発します (Ramón y Cajal. 1898)。古くはHippocratesも、左頭部負傷が右の筋群の運動に影響を与えることを知っていたようです (Vulliemoz et al., Lancet Neurol. 2005)。しかしながら、神経科学の著名な教科書にもこれに対する明確な回答はありません (カンデル神経科学や神経科学-脳の探求-を一通りざっと見ただけですが)。幸い、英語でGoogle検索すると当然ながら同様の疑問を抱く人は多数いるようで、Quoraにおいていくつかの解答を見つけました (Why does our left hemisphere of brain control our right side of our body and the right our left? - Quora)。
【追記(2020/03/10)】WikipediaのContralateral brainのページとも内容が被っていることに気づきました。
Quoraの記事で紹介されている論文を足掛かりに文献を調べた結果ですが、この問題に関する優れた総説として(Vulliemoz et al., Lancet Neurol. 2005; Mora et al., Neurosurg. Focus. 2019)があります。
本記事では、対側支配の神経解剖を復習しつつ、脳が対側支配をする進化的な利点の仮説を紹介していきます。なお、仮説ということを強調しているのはこれが進化的な話を含み、実験的証明が難しいためです。また、筆者は神経解剖学・神経発生学の教室でも研究をしていますが、本記事における内容に関しては全く研究していないのでご了承ください。
正中を横切る神経線維
まず、正中を横切る神経線維は交叉(decussation)や交連(commissure)と言った名称がつけられています。ここではいくつかの経路を簡単にまとめましたが、詳しくは何処のご家庭にもある神経解剖の教科書を読みましょう (今回は寺島先生の神経解剖学講義ノートを参考にしました)。
視交叉(optic chiasma)
視神経の交叉のことです。両生類、魚類、鳥類では視神経が完全に交叉します(完全交叉)が、ネコなどの食肉類や霊長類では50%の視神経のみが交叉します(半交叉)。
運動路における交叉
運動路で代表的なのが皮質脊髄路(corticospinal tract, CST; または錐体路とも)です。皮質脊髄路は主に運動性皮質L5の錐体細胞を起始とし、脊髄に終わる経路です。皮質脊髄路の途中で延髄腹側の錐体において神経線維が交叉しますが、これを錐体交叉(または運動交叉) (pyramidal decussation, motor decussation)と言います。錐体交叉では全ての神経が交叉するわけではないですが、交叉しなかった線維も脊髄の白交連(anterior white commissure)で最終的には交叉します。なので、皮質脊髄路線維は全て対側の運動ニューロンを支配することになります。他には赤核脊髄路 (rubrospinal tract)における腹側被蓋交叉(tegmental decussation)や、視蓋脊髄路 (tectospinal tract)における背側被蓋交叉(dorsal tegmental decussation)があります。
感覚路における交叉
感覚路も色々ありますが、頭部以外の体性感覚の伝導路であれば後索・内側毛帯系と脊髄視床路系があります。後索・内側毛帯系では毛帯交叉(sensory decussation, decussation of the lemniscus)、脊髄視床路系では脊髄の白交連を経由して対側の体性感覚野に至ります。
小脳の経路における交叉
念のため触れておきます。まず、小脳は同側支配です。例えば右小脳半球外側部を損傷すると、右側 (患側)に小脳性運動失調が現れます。この理由は、小脳からの出力線維が交叉しない(例えば非交叉性室頂核前庭線維)か、2回交叉するためです。2回交叉は裏の裏が表になるのと同じことで、例えば上記の症例における経路は、小脳半球外側部から視床VL核までで上小脳脚交叉があり、視床から大脳皮質は交叉無し、大脳皮質から皮質脊髄路の錐体交叉でもう一度交叉する、となっています。
神経線維の交叉に関連した疾患
神経系の発生においては軸索誘導分子(axon guidance molecules)の寄与が必要ですが、それらをコードする遺伝子に異常が生じると神経発生が正常に行われなくなります。そのような疾患の例として、クリッペル・ファイル症候群(Klippel-Feil syndrome)、 X連鎖性カルマン症候群(X-linked Kallmann’s syndrome)などでは非交叉性の皮質遠心性線維が形成されることで鏡像運動(mirror movements; 随意的な運動を行うときに、対側にも不随意的に運動が生じる症状)が生じます (Comer et al., Neural Dev. 2019)。しかしながら病態機序はいくつか考えられるようなので、詳細は書きません (cf. 脳科学辞典の鏡像運動の項目)。
Ramón y Cajalの仮説
この節は、Ramón y Cajalの仮説についての説明をします。なお、Cajalの発表の後となりますが英国の医師であったFrancis DixonもCajalと類似の説を提唱しています(Dixon, The Dublin Journal of Medical Science. 1907; Dixon, The Dublin Journal of Medical Science. 1918)。
さて、Cajalの視交叉についての仮説は、脳内での外界の連続性を維持するためには交叉が必要である、ということです。次図は前方に眼があり視交叉が無い生物が矢を見た場合の図です。網膜で外界が反転されるため、交叉が無いと矢尻と矢羽が繋がった像となります。そのため、2つの視野像から脳内で外界を復元するには像を交叉させる必要があります。
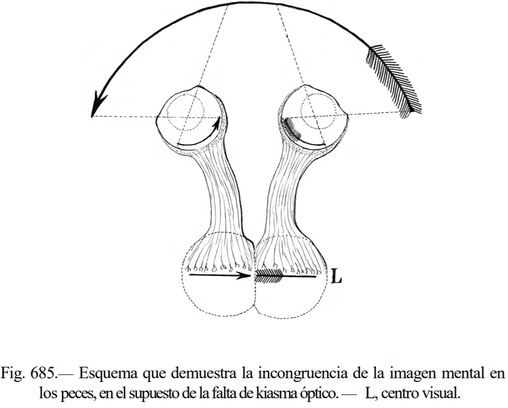
(Ramón y Cajal. 1898; 図の間接的な引用元は Ramón Y Cajal. Histología del sistema nervioso del hombre y de los vertebrados. Tomo II. Segunda parte. 2012)
次図の上部は、前述の通り視交叉により外界が正しく復元されることを示しています。また、完全な交叉ではない半交叉であることが立体視に重要であることも指摘していました。Cajalはさらに説を広げ、運動路と感覚路に交叉がある理由も視交叉と関連付けました。これが対側支配におけるCajalの仮説です。次図中央部では連続した外界が復元されていますが、このままでは像が反転しています。Cajalは視野の右側を体の右側と、視野の左側を体の左側と一致させるために、運動路と体性感覚路を交叉させる必要があると主張しました。
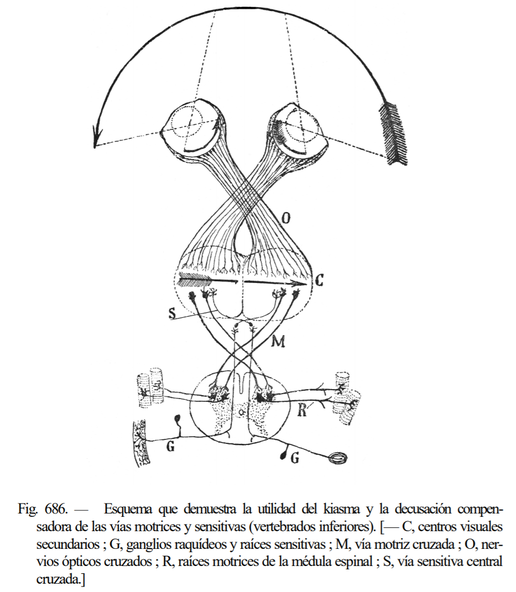
しかし、Cajalの仮説は完全に誤っているわけではないものの、いくつかの問題点があることが指摘されています (de Lussanet & Osse, Animal Biol. 2012)。まず、視交叉の説について、眼球は動くため、網膜像も動きます。そのため、位置合わせを完全に正確にすることはできません (予測により補間はしていると思われるので、Cajalが間違っているとは言えませんが)。次に、運動路と感覚路を関連させた説については、同側の視覚情報・感覚情報のみが運動野に至るというのは最適ではないということです。運動することを考えれば、両側の情報を統合して座標系を生み出した方がよいでしょうし、実際に脳はそうしています。
ということで、Cajalの仮説は完全に誤りではないにせよ、間違っている点も見られるということになります。ただ、Cajalが偉大だったことに変わりはないなと、今回スケッチを見直して思いました。
ねじれ仮説 (twist hypothesis)
無脊椎動物から脊椎動物へ至る間に体のねじれが生じた、というのがねじれ仮説 (twist hypothesis) です。これは対側支配が生じた目的ではなく、その過程についての仮説となっています。
ねじれ仮説には(Kinsbourne, Neuropsychology. 2013)の体性ねじれ説 (somatic twist hypothesis) と、(de Lussanet & Osse, Animal Biol. 2012)の軸ねじれ説 (axial twist hypothesis) の2つが提案されています。まず、Kinsbourneの体性ねじれ説は、180度の回転が一回起こった、というものです。一方で、de LussanetとOsseの軸ねじれ説は2つの90度の回転が起こった、というものです。両者の差異は分かりづらいですが、de LussanetとOsseによって解説されています (de Lussanet & Osse, Neuropsychology. 2015)。
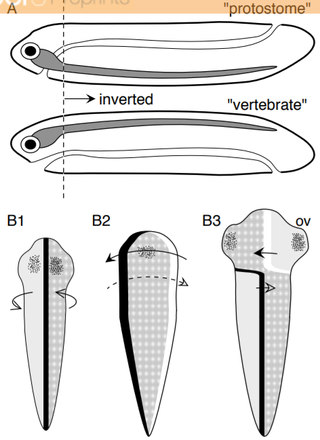
AはKinsbourneの体性ねじれ説を示しており、前口動物(Protostome)が脊椎動物(vertebrate)へと進化するに伴い、前脳と眼の領域が180度回転したことを表します。Bはde LussanetとOsseの軸ねじれ説で、黒線が背側、白線が腹側、点は眼となる領域、ovはoptic vesicleを意味します。重要なのはB2で、前脳と眼の領域が90度の時計回り、中脳より下が90度の反時計回りをすることで合計で180度のねじれが生まれます。
どちらの仮説も視交叉や、交叉しない嗅覚経路、および小脳の同側支配などを説明します。しかし、de LussanetとOsseは軸ねじれ説であれば、心臓や腸管などの臓器が左右対称ではないこと、などが説明できると主張しています。また、両者の説の弱いところとして視交叉などでの全交叉は説明しても、一部の神経が同側に向かう半交叉は説明できない、といった点を挙げています。
いずれにせよ、進化の過程で何らかのねじれが生じたことは誤りではないでしょうが、何故そのねじれが保存されてきたのでしょうか?やはり、そこには対側支配の利点があったのでしょう。次節からは対側支配(主に皮質脊髄路や赤核脊髄路の運動制御、脊髄視床路の体性感覚)の進化的利点についての仮説を見ていきます。
回避行動仮説 (avoidance behavior hypothesis)
危険な刺激を知覚し回避行動を取るために体の構造の進化に伴って交叉が必要となった、というのが回避行動仮説 (avoidance behavior hypothesis) です (Vulliemoz et al., Lancet Neurol. 2005)。
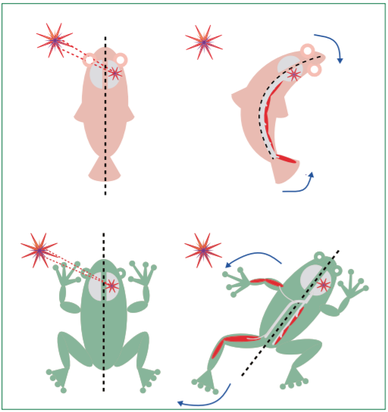
まず、交叉が無く、手足も無い原始的な種の場合を考えます(図上部; なお図では魚のような見た目ですが、魚類の錐体路も交叉が存在するようです)。このような生物の場合、危険な刺激は右脳半球によって知覚され、右半身の筋肉の収縮を引き起こし、屈曲させることで危険を回避します。このとき、交叉していない原始的な経路 (網様体脊髄路や前庭脊髄路)を介して筋肉は制御されます。
一方で、四肢を持つ脊椎動物(図下部)は、左肢を伸ばすことにより、左側の刺激を回避しようとします。このとき、交叉のある(系統発生的に)新しい経路 (皮質脊髄路や赤核脊髄路)を介して筋肉は制御されます。
特に根拠についての記載はありませんでしたが、1つの説と言えます。
保護機構仮説 (protective mechanism hypothesis)
対側支配であることは身体保護の観点で優れている、という説が保護機構仮説 (protective mechanism hypothesis) です (Whitehead & Banihani, Laterality. 2014)。前節の回避行動仮説と体を守るという観点では似ていますが、少し異なります。これまでに紹介してきた論文と異なり、WhiteheadとBanihaniは中程度の損傷を受けた場合、対側支配の方が生存しやすいことを簡単な数理モデルを用いて説明しています。ここでは数理モデルの紹介は省略し、簡単なお気持ちだけ説明します。
ここでは、生物の体が左右の脳半球(と神経)及び左右の筋群の4つから成ると考えます。両半身を損傷すると致命的となることが多いですが、片半身だけの損傷であれば生存することも可能です。このとき、例えば右脳半球と左筋群の組み合わせが損傷を受けることはあまりなく、どちらかというと転倒や衝突により片側の脳半球と筋群が傷害されやすいでしょう。
同側支配の場合、片側の脳半球と筋群が同時に損傷を受けると、損傷を受けた半球と損傷を受けた筋群という組み合わせにより片側は完全に動かすことができなくなります。一方で対側支配の場合、各脳半球は反対側を制御するため、例えば右半身を損傷した場合は、損傷した右半身を制御する無傷の左脳半球と、無傷の左側を制御する損傷した右脳半球という状況になります。無傷の左半球は損傷しているとは言え、一部の右側の筋群を動かすことができるでしょう。また、損傷した右半球であっても左側の筋群は無事であるためにある程度は制御できるでしょう。
片側が完全に機能しない場合に比べ、両側に影響はあるものの何とか動かすことができる場合の方が生存しやすいと仮定すると、交叉が保存されてきたことにも納得できます。
まとめ
本記事では主に運動野と感覚野の対側支配がどのように生じたのか、また何故対側支配が保存されてきたのか、ということについての仮説を見てきました (重ねて言いますが仮説止まりです)。現在提案されている説をまとめると、体がねじれることで対側支配が生まれ、それが生存にとって有利であったために保存されてきた、と言えます。冒頭に述べたように、進化が絡むと直接的な証拠というものを見つけづらいとは思いますが (他の研究例を知らないだけかもしれませんが)、将来は何らかの説に落ち着くのではないか、と思っています。
参考文献
- Carson RG. Neural pathways mediating bilateral interactions between the upper limbs. Brain Res Rev. 2005;49(3):641–662. doi:10.1016/j.brainresrev.2005.03.005
- Comer JD, Alvarez S, Butler SJ, Kaltschmidt JA. Commissural axon guidance in the developing spinal cord: from Cajal to the present day. Neural Dev. 2019;14(1):9. doi:10.1186/s13064-019-0133-1
- de Lussanet MH, Osse JW. Decussation as an axial twist: A comment on Kinsbourne (2013). Neuropsychology. 2015;29(5):713–714. doi:10.1037/neu0000163
- de Lussanet, MH, Osse, JW. An ancestral axial twist explains the contralateral forebrain and the optic chiasm in vertebrates. Animal Biol. 2012; 62: 193-216. doi:10.1163/157075611X617102.
- Dixon AF. Why are the great motor and sensory tracts of the central nervous system crossed?. The Dublin Journal of Medical Science. 1907; 124, 1–4. doi:10.1007/BF02972358
- Dixon AF. Why are the cerebral motor and sensory cortical areas arranged in an inverted order?. The Dublin Journal of Medical Science. 1918; 145, 154–160. doi:10.1007/BF02958527
- Kinsbourne M. Somatic twist: a model for the evolution of decussation. Neuropsychology. 2013;27(5):511–515. doi:10.1037/a0033662
- Loosemore RG. The inversion hypothesis: A novel explanation for the contralaterality of the human brain. Biosci Hypotheses. 2009;2:375–382
- Mora C, Velásquez C, Martino J. The neural pathway midline crossing theory: a historical analysis of Santiago Rámon y Cajal’s contribution on cerebral localization and on contralateral forebrain organization. Neurosurg Focus. 2019;47(3):E10. doi:10.3171/2019.6.FOCUS19341
- Ramón y Cajal, S. Estructura del kiasma óptico y teoría general de los entrecruzamientos de las vías nerviosas. 1898. [german 1899, english 2004]. Rev. Trim. Microgràfica 3, 15–65.
- Ramón Y Cajal, S. Histología del sistema nervioso del hombre y de los vertebrados. Tomo II. Segunda parte. Boletín Oficial del Estado. 2012.
- Shinbrot T, Young W. Why decussate? Topological constraints on 3D wiring. Anat Rec (Hoboken). 2008;291(10):1278–1292. doi:10.1002/ar.20731
- Vulliemoz S, Raineteau O, Jabaudon D. Reaching beyond the midline: why are human brains cross wired?. Lancet Neurol. 2005;4(2):87–99. doi:10.1016/S1474-4422(05)00990-7
- Welniarz Q, Dusart I, Roze E. The corticospinal tract: Evolution, development, and human disorders. Dev Neurobiol. 2017;77(7):810–829. doi:10.1002/dneu.22455
- Whitehead L, Banihani S. The evolution of contralateral control of the body by the brain: is it a protective mechanism?. Laterality. 2014;19(3):325–339. doi:10.1080/1357650X.2013.824461